Sexual versus asexual reproduction
Meiosis
Genetic recombination by crossing over
Determination of sex
Sexual versus asexual reproduction
Sexual reproduction has a few disadvantages compared to asexual reproduction. It requires more than just one individual to parent each offspring, rendering about half of the individuals unable to carry through the bearing of offspring, while allowing just half of each parent’s contribution to be inherited by their offspring.
The advantage that makes it significant enough to outweigh the disadvantages is increased diversity. With each generation, diversity is inevitably inscribed into offspring, and the sources of genetic variation maximised with each cross. In sexual reproduction, individuals are simply forced to mix by default. As such, even very accomplished parental genomes are inevitably disrupted.
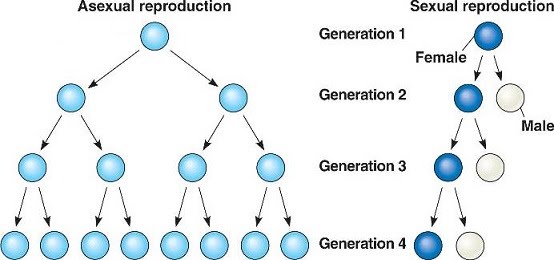
The overall effect is a better position for responding to selection pressures, as exemplified by the Red Queen hypothesis where individuals rely on varied strategies to keep up with their parasites, predators, etc. Since the standard for fitness is ever-changing, a real time diversified gene pool is maintained in populations.
Asexual reproduction is optimal in niche environments that are more stable, or when recolonising habitats that were previously disturbed. Good example of this are plants that reproduce clonally, and parthenogenic animals. It is advantageous for plants to spread via any part of their body e.g. stem, roots, rather than have to rely on fertilisation via flowers. This ability is used by growers to study cloned plants.
Parthenogenic animals reproduce asexually using unfertilised eggs. In cooler environments, the pressures from parasites are lower, as their density is lower and they cannot thrive in the cold. This allows parthenogenic animals to reproduce asexually and forego the higher diversity enabled by sexual reproduction.
Some asexually reproducing organisms such as bacteria have options for horizontal transfer of genetic information e.g. the sharing of plasmids between individuals, even between different species.
Meiosis
Meiosis is a type of cell division which results in 4 cells that are genetically non-identical from one parent cell. In order for once cell to divide to result in 4 cells, how many divisions must take place?
Two. 1 cell becomes 2, then 2 become 4:
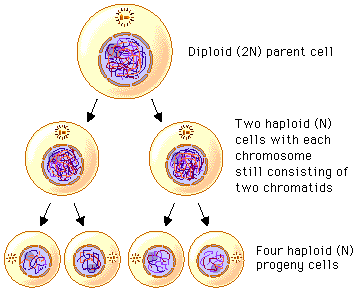
The first division is called meiosis I, and the second is called meiosis II.
…so far so easy? (it should be!)
Cells resulting from meiosis are gametes such as egg cells and sperm cells, hence meiosis only occurs in sexually reproducing organisms. There are 2 key points about this…