Introduction
How does water enter a plant?
Xylem
The vascular system of a plant
The 3 pathways that water uses
Transpiration
Root Pressure, The Cohesion-Tension hypothesis
Hypothesis of sugar movement through phloem
Introduction
Mass transport is required in multicellular plants due to their increased size, the variation in their metabolism as they photosynthesise and respire at different rates throughout the day or in different seasons, and the decreased surface area to volume ratio coming from their larger size.
How does water enter a plant?
Water enters a plant through the roots. In order to understand how water gets in the root, you should definitely check out the root structure:
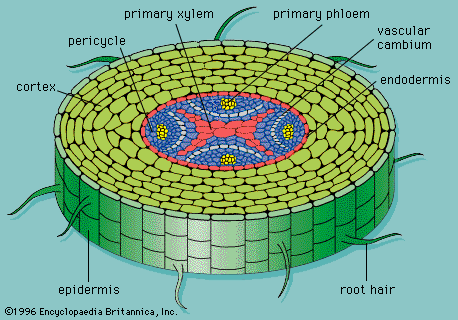
What you can see above is a delicious slice of pineapple. OK, it’s not. That is a slice of a root. Roots, as you may have seen in real life, are hairy. All those tiny and not so tiny root hairs buried into the soil greatly increase the surface area of the root. This exposes it to more water molecules which can be taken up. The hairs are nothing like human hairs; they are extensions of the outer layer of the root, made up of cells. This layer is called the epidermis.