Detecting and Amplifying and environmental Stimulus
Photoreceptors
Vision Photoreceptors
How?
Introduction
Photoreceptors are an example of receptors found across all three domains of life. In addition to the ubiquitous use in the animal kingdom of eyes, and light-using photosynthesis in plants, photoreceptors are also central to the driving force of proton gradients in archaea for their cellular functions.
Some archaea use light to drive this process, while others use vastly diverse sources of energy e.g. metal ions.
Cellular respiration produces ATP (adenosine triphosphate), the energy currency in biochemical processes. Oxygen-aided respiration – aerobic respiration – produces much more ATP than anaerobic respiration.
ATP is made through ADP by adding back its third phosphate, and it reacts reversibly by losing that phosphate and releasing energy.
We are talking about adenosine triphosphate breaking down into adenosine diphosphate, inorganic phosphate and energy; and the latter joining back together to make adenosine triphosphate again.
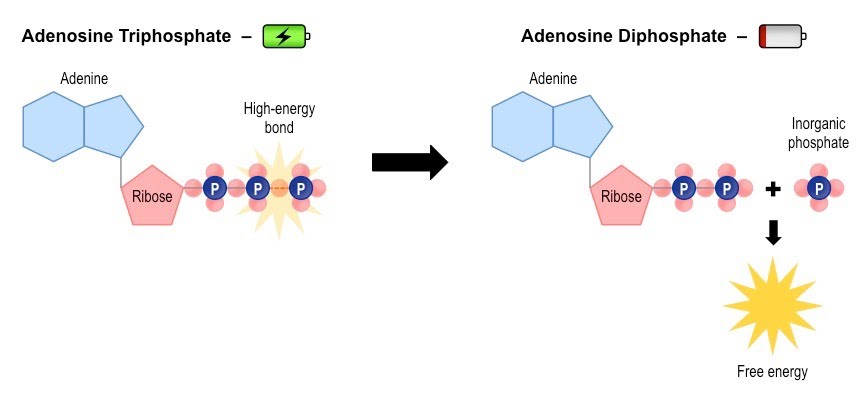
When the hydrolysis of ATP (via the enzyme ATP hydrolase) is coupled to other reactions requiring energy, it enables these processes to take place. The inorganic phosphate released can itself take part in a further phosphorylation reaction with another chemical, often increasing its reactivity.
The condensation of ADP and inorganic phosphate takes place during photosynthesis and respiration, and it is catalysed by the enzyme ATP synthase. Because it synthesises ATP. Get it get it..
ATP synthase (also known as ATP synthetase) produces ATP when hydrogen ions, H+, pass through its transmembrane channel. The reason they pass through is that there are more of them on one side of the semipermeable membrane than the other. This movement is called chemiosmosis.
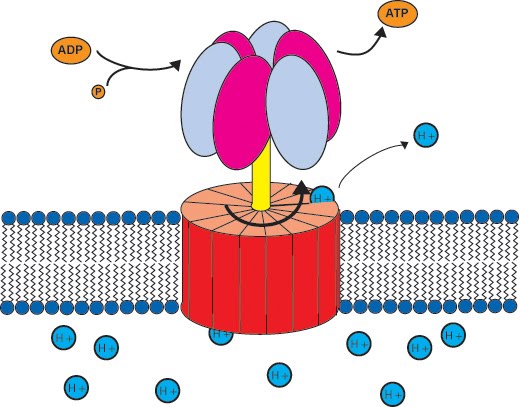
As protons (H+) are driven to cross the membrane via ATP synthase, this rotates and catalyses one ADP and one…..