Auxins and cytokinins
Gibberellins and phytochromes
Auxins and cytokinins
Plants, too, have hormones. Ooooooooooooooooooh. They control plant growth such as whether they grow to the left or to the right in response to light. Some key hormones are auxin (more specifically indoleacetic acid/IAA), cytokinins and gibberellins. Gibberellin is a fun word, just say it, geeee-berrrrrrr-elllllllll-eeeeen-brbrlrbrlrbbbrlrlrllllllllllll.
Auxin stimulates cell elongation in the roots and shoots in flowering plants. It accumulates away from light which simulates growth on the dark side, bending the shoot in the opposite direction, where the light is.
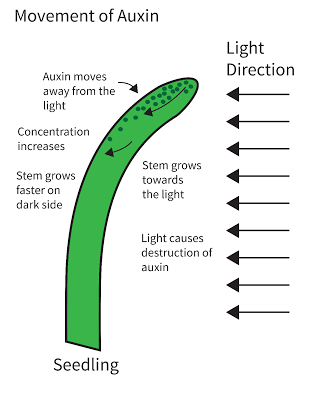
This ensures plants respond to their environment in a way that best maximises their survival prospects. More light enables better photosynthesis, while growing upwards exposes their leaves to more sunlight. Auxin also suppresses lateral bud growth, so only the apical bud is growing, as it secretes auxin that travels downwards. This is called apical dominance and it ensures that the shoot grows tall rather than wide. If the apical bud is removed lateral buds sprout.
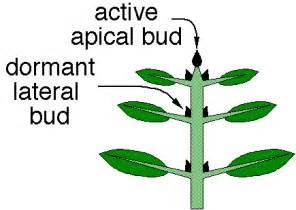
Cytokinins are made in the roots and travel upwards to promote lateral plant growth. In this way, they’re working opposite to auxin. Overall they strike a balance to drive good growth..