Photosynthesis
Light-dependant and light-independent reactions
Limiting factors
The Compensation Point
Microorganisms in the Ecosystem
Biomass Transfer through the food chain
Farming and the Environment
Abiotic factors
Biotic Factors
Practices that boost food production at the expense of Biodiversity
Leaching and Eutrophication
Pollution by slurry and silage effluent
Promoting biodiversity
Polyculture
Succession
Field investigation of Biodiversity
Recording data
Photosynthesis
Photosynthesis is a metabolic process which makes stuff using light. How? How can you make anything from light? And why? Living things are made of complex organic molecules such as carbohydrates and proteins, as opposed to simple inorganic molecules such as carbon dioxide and water.
The vast majority of plants on Earth today undergo photosynthesis via a specific route (C3) which is slightly different to two other potential routes (C4 and CAM). The general balanced reaction for photosynthesis is:
H2O + CO2 + energy –> C6H12O6 + O2
…where water, carbon dioxide and energy are the starting materials, and glucose and oxygen the products. Here, glucose is the key product because it is the complex organic molecule made from simple inorganic reactants. The “energy”, as you may have noticed, is where the light comes in.
The energy stored in big molecules (such as carbohydrates) created via photosynthesis is derived in part through the light energy in photons. In order to tap into this energy, light must be absorbed by plants and other photosynthetic organisms.
As you know, visible light ranges in wavelength with colour:
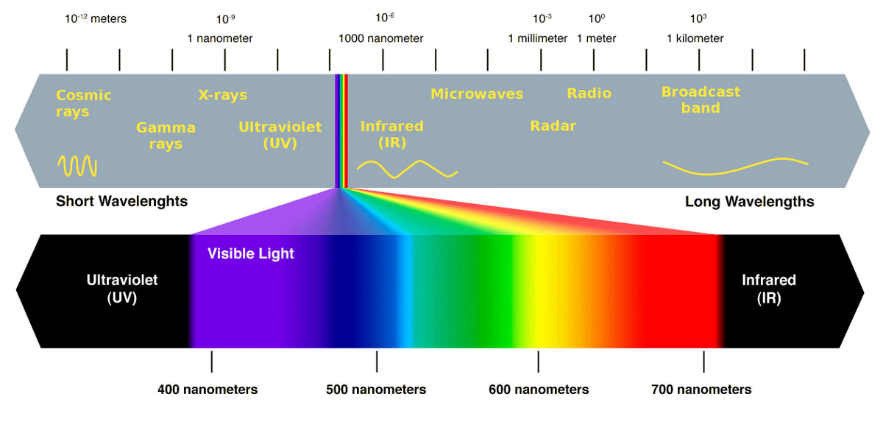
Between 400-700 nm, light passes through several colours from violet to red. Pigments absorb some wavelengths more than others, just like anything else we see as coloured. For example, something appears yellow if it absorbs other colours like blue (500 nm) and red (700 nm) but reflects yellow (600 nm).
The different pigments can be extracted from a plant by grinding its tissue in a solvent which will become green…..