Parental investment
Courtship
Sexual reproduction brings with it its own set of behavioural implications, as individuals in a species must interact for the purpose of reproduction.
Parental investment
Parental investment refers to the input required by parents to ensure the survival of their offspring. This can be diverged even before the offspring are brought to life, in terms of the preparations required, whether biological or environmental. For example, the production and incubation of eggs requires more investment of energy for fewer offspring, while the production of sperm cells requires less energy for more cells.
In organisms that are immobile i.e. sessile, such as barnacles, sex poses a distinct problem.
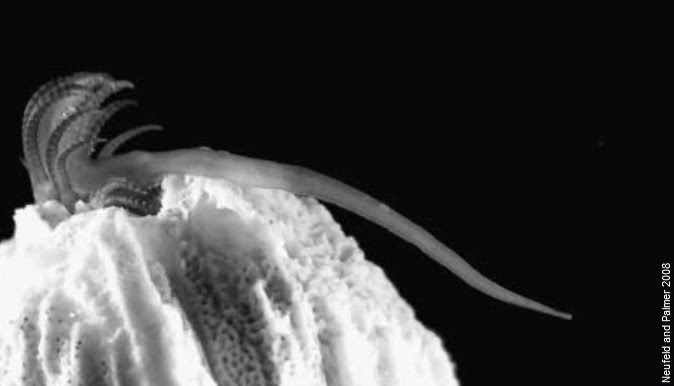
Barnacles are attached to their rock, and cannot approach one another for sexual reproduction. A solution for this is the very long barnacle penis that extends quite some distance from its body in the hope of reaching a receptive barnacle nearby. Another solution is the casting of sperm into the sea for other barnacles to passively pick up.
This highlights internal versus external fertilisation. Each has its own costs and benefits. External fertilisation such as that carried out by fish requires little energy in terms of finding a mate and protecting offspring, and can produce many offspring over a large area, but results in higher mortality of offspring and wasted gametes.
Internal fertilisation on the other hand has better offspring survival as the embryo is shielded away from predators and offspring taken care of by parents, at the expense of having fewer offspring and expending energy towards finding a mate and looking after offspring.
In r/K selection theory, the number of offspring a population can have in a habitat is used to contrast species that produce many offspring with those that produce few.

r refers to the rate of growth while K refers to the carrying capacity of the habitat. Species that invest less in their offspring but have many are on the r side of the equation, hence termed r-selected organisms.
Those that tend towards the K side have fewer offspring and invest more energy in each of them. These are K-selected organisms.
Several traits are associated with each reproductive strategy, such as organism size, the stability of the niche they occupy and the developmental speed of offspring. Large organisms have fewer offspring and take close care of them. The offspring reach maturity slower….