Introduction
Transpiration
Root pressure. The cohesion-tension hypothesis
Arguing the mass-flow hypothesis of sugar movement through phloem
Introduction
Water enters a plant through the roots. In order to understand how water gets in the root, you should definitely check out the root structure:
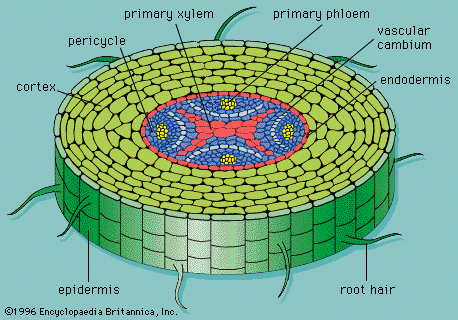
What you can see above is a delicious slice of pineapple. OK, it’s not. That is a slice of a root. Roots, as you may have seen in real life, are hairy. All those tiny and not so tiny root hairs buried into the soil greatly increase the surface area of the root. This exposes it to more water molecules which can be taken up. The hairs are nothing like human hairs; they are extensions of the outer layer of the root, made up of cells. This layer is called the epidermis.
Why does water move inside the root? Simple: osmosis. The cell sap (i.e. cell juice) has a lower water potential than the fluid found in the soil, so the water in the soil kindly makes its way into the thirsty awaiting root. Once the water reaches the first cell in its path, the water potential of that cell is increased compared to the cell next to it. Therefore, water moves into the next cell, leaving the current cell. This in turn results in the previous cell taking up water all over again, and so forth, until the water makes its way across all cells of the cortex.
Reaching the endodermis, water then enters the xylem. The xylem is a tissue of dead cells which contributes to the vascular system of plants by being the transportation medium for water and dissolved mineral ions. The xylem brings them to the leaves and plants’ other organs.
There are two different pathways that water uses in order to reach the xylem:
1. The apoplast pathway whereby water slaloms between cell walls and the spaces in between, without passing directly through live tissue; this accounts for 90% of water uptake.
2. The symplast pathway whereby water goes straight through living tissue i.e. the cells in the cortex, and into the xylem; this accounts for only 10% of water uptake.
Basically, the symplast pathway is just way simpler.
Transpiration
Transpiration is water loss through the parts of a plant which are found above soil level i.e. not the roots. As water streams through a plant, transpiration affects the speed of the stream. Increased transpiration will lead to a quicker uptake of water through the roots to maintain the water flow throughout the plant. So what affects transpiration?
1. Light causes stomata to open, resulting in increased water loss (transpiration).
2. Temperature going up also raises the rate of transpiration, as more water molecules evaporate.
3. Humidity. An increase in humidity around the leaves means that there is less space for water molecules from the plant to evaporate into, so transpiration is decreased.
4. Air movement (wind) can displace water molecules from around the stomata, so that more space becomes available for additional water molecules to go into. Transpiration increases.
Root pressure. The cohesion-tension hypothesis
These are the two ways in which the stream of water through a plant can work. Root pressure is the water being pushed into the roots, while cohesion-tension is the water being pulled up…